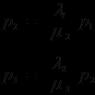Примеры цепи и сети питания. Пищевые цепи и пищевые сети
При изучении биотической структуры экосистемы становится очевидным, что одно из важнейших взаимоотношений между организмами - это пищевое. Можно проследить бесчисленные пути движения вещества в экосистеме, при котором один организм поедается другим, а тот - третьим и т.д.
Детритофаги
Орёл Детритофаги V
Лиса Человек Орёл Детритофаги IV
Мышь Заяц Корова Человек Детритофаги III
Пшеница Трава Яблоня I
Пищевая цепь - это путь движения вещества (источник энергии и строительный материал) в экосистеме от одного организма к другому.
Растение корова
Растение корова человек
Растение кузнечик мышь лиса орёл
Растение жук лягушка змея птица
Обозначает направление движения.
В природе пищевые цепи редко изолированы друг от друга. Гораздо чаще представители одного вида (растительноядные) питаются несколькими видами растений, а сами служат пищей для нескольких видов хищников. Перенос вредных веществ в экосистеме.
Пищевая сеть - это сложная сеть пищевых взаимоотношений.
Несмотря на многообразие пищевых сетей, они все соответствуют общей схеме: от зелёных растений к первичным консументам, от них к вторичным консументам и т.д. и к детритофагам. На последнем месте всегда стоят детритофаги, они замыкают пищевую цепь.
Трофический уровень - это совокупность организмов, занимающих определённое место в пищевой сети.
I трофический уровень - всегда растения,
II трофический уровень - первичные консументы
III трофический уровень - вторичные консументы и т.д.
Детритофаги могут находиться на II и выше трофическом уровне.
III 3,5 дж вторичный консумент (волк)
II 500 дж первичный консумент (корова)
I 6200 дж растения
2,6*10 дж поглощено солнечной энергии
1,3*10 дж падает на поверхность земли на
некоторую площадь
Пирамида энергии
III 10 кг лиса (1 )
II 100 кг заяц (10 )
I 1000 кг растения на лугу (100 )
Пирамида биомассы.
Обычно в экосистеме насчитывается 3-4 трофических уровня. Это объясняется тем, что значительная часть потребляемой пищи тратится на энергию (90 - 99 %), поэтому масса каждого трофического уровня меньше предыдущего. На формирование тела организма идет относительно немного (1 - 10 %.Соотношение между растениями, консументами, детритофагами выражают в виде пирамид.
Пирамида биомассы - показывает соотношение биомасс различных организмов на трофических уровнях.
Пирамида энергии- показывает поток энергии через экосистему. (см.рис.)
Очевидно, что существование большего числа трофических уровней невозможно, из-за быстрого приближения биомассы к нулю.
Автотрофы и гетеротрофы.
Автотрофы - это организмы, способные строить свои тела за счет неорганических соединений, используя солнечную энергию.
К ним относятся растения (только растения). Они синтезируют из СО , Н О (неорганические молекулы) под воздействием солнечной энергии - глюкозу (органические молекулы) и О . Они составляют первое звено в пищевой цепи и находятся на 1 трофическом уровне.
Гетсротрофы - это организмы, которые не могут строить собственное тело из неорганических соединений, а вынуждены использовать созданное автотрофами, употребляя их в пищу.
К ним относятся консументы и детритофаги. И находятся на II и выше трофическом уровне. Человек тоже гетеротроф.
Вернадскому принадлежит идея, что возможно превращение человеческого общества из гетеротрофного и автотрофное. В силу своих биологических особенностей человек не может перейти к автотрофности, но общество в целом способно осуществить автотрофный способ производства пищи, т.е. замена природных соединений (белки, жиры, углеводы) на органические соединения, синтезированные из неорганических молекул или атомов.
Любой живой организм выбирает условия, наиболее благоприятные для его обитания и обеспечивающие его возможностью полноценно питаться. Лиса выбирает место проживания, где живет много зайцев. Лев селится поближе к стадам антилоп. Рыба-прилипала не только путешествует, прикрепившись к акуле, но и сотрапезничает с ней.
Растения, хотя и лишены возможности сознательно выбрать место обитания, но преимущественно произрастают тоже в наиболее комфортных для себя местах. Серую ольху часто сопровождает крапива, требовательная к азотному питанию. Дело в том, что ольха сожительствует с бактериями, обогащающими почву азотом.
Пищевая сеть - своеобразный симбиоз
Здесь мы сталкиваемся с определенным видом отношений. Речь идет о так называемом симбиозе. Это прямые отношения, при которых оба организма извлекают пользу. Их также называют пищевые сети и цепи. Оба термина имеют похожее значение.
Чем отличаются друг от друга пищевая цепь и пищевая сеть? Отдельные группы организмов (грибы, растения, бактерии, звери) постоянно обмениваются между собой определенными веществами и энергией. Данный процесс называется пищевой цепью. Обмен между группами осуществляется во время поедания одних другими. Процесс взаимодействия между подобными цепями называется пищевой сетью.
Как организмы взаимосвязаны
Известно, что бобовые растения (клевер, мышиный горошек, караганы) сожительствуют с клубненьковыми бактериями, переводящими азот в формы, которые усваиваются растениями. В свою очередь бактерии получают от растений необходимые им органические вещества.
Многие из описанных отношений носят специфический характер. Однако в каждом биоценозе существуют отношения, в которых принимает участие каждая популяция. Это пищевые или трофические (трофос - пища) отношения.

Примеры пищевых сетей и цепей:
Во всех случаях организм, питающийся другими, извлекает одностороннюю пользу. Участвуя в процессе питания, все особи популяции обеспечивают себя необходимой для их жизнедеятельности энергией и разнообразными веществами. Популяция, служащая объектом питания, испытывает отрицательное воздействие со стороны пожирающих ее хищников.
Аутотрофы и гетеротрофы
Вспомним, что по способам питания организмы делятся на две группы.
Аутотрофные (аутос - сам) организмы живут за счет неорганического источника углеводорода. В данную группу входят растения.

Гетеротрофные (гетерос - другой) организмы живут за счет органического источника углеводорода. В эту группу входят грибы, бактерии. Если автотрофы независимы от иных организмов в источнике углерода и энергии, то гетеротрофы в этом отношении полностью зависят от растений.
Конкурентные отношения между группами
Взаимоотношения, приводящие к угнетению одного из партнеров, не обязательно связаны отношениями питания. Многие сорняки выделяют метаболиты, задерживающие рост растений. Одуванчик, пырей ползучий, василек угнетающе действуют на овес, рожь и другие культивируемые злаки.
В каждом биоценозе живут популяции многих видов, и отношения между ними многообразны. Можно сказать, что популяция ограничена в своих возможностях этими отношениями и должна найти присущее только ей место.

Уровень обеспеченности места обитания экологическими ресурсами определяет возможность существования многих ниш. От этого зависит и число видовых популяций, образующих биоценоз. В условиях благоприятного климата степей формируются биоценозы, состоящие из сотен видов, а в тропическом климате леса - из тысячи видов организмов. Пустынные биоценозы в жарком климате насчитывают несколько десятков видов.
Столь же изменчиво и пространственное распределение популяций. Тропические леса многоярусны, и живые организмы заполняют весь объем пространства. В пустынях биоценозы просты по структуре, и популяции малочисленны. Таким образом видно, что совместная жизнь организмов в биоценозах необыкновенно сложна. И все-таки растения и звери, грибы и бактерии объединяются в биоценозы и существуют только в их составе. Каковы же причины этого?
Самой главной из них является потребность живых организмов в питании, в трофической зависимости друг от друга.
В природе любой вид, популяция и даже отдельная особь живут не изолированно друг от друга и среды своего обитания, а, напротив, испытывают многочисленные взаимные влияния. Биотические сообщества или биоценозы - сообщества взаимодействующих живых организмов, представляющие собой устойчивую систему, связанную многочисленными внутренними связями, с относительно постоянной структурой и взаимообусловленным набором видов.
Для биоценоза характерны определенные структуры : видовая, пространственная и трофическая.
Органические компоненты биоценоза неразрывно связаны с неорганическими - почвой, влагой, атмосферой, образуя вместе с ними устойчивую экосистему - биогеоценоз .
Биогеноценоз – саморегулирующаяся экологическая система, образованная совместно обитающими и взаимодействующими между собой и с неживой природой, популяциями разных видов в относительно однородных условиях среды.
Экологические системы
Функциональные системы, включающие в себя сообщества живых организмов разных видов и их среду обитания. Связи между компонентами экосистемы возникают, прежде всего, на основе пищевых взаимоотношений и способов получения энергии.
Экосистема
Совокупность видов растений, животных, грибов, микроорганизмов, взаимодействующих между собой и с окружающей средой таким образом, что такое сообщество может сохраняться и функционировать необозримо длительное время. Биотическое сообщество (биоценоз) состоит из сообщества растений (фитоценоз ), животных (зооценоз ), микроорганизмов (микробоценоз ).
Все организмы Земли и среда их обитания также представляют собой экосистему высшего ранга - биосферу , обладающую устойчивостью и другими свойствами экосистемы.
Существование экосистемы возможно благодаря постоянному притоку энергии извне - таким источником энергии, как правило, является солнце, хотя не для всех экосистем это справедливо. Устойчивость экосистемы обеспечивается прямыми и обратными связями между ее компонентами, внутренним круговоротом веществ и участием в глобальных круговоротах.
Учение о биогеоценозах разработано В.Н. Сукачевым. Термин «экосистема » введен в употребление английским геоботаником А. Тенсли в 1935 г., термин «биогеоценоз » - академиком В.Н. Сукачевым в 1942 г. В биогеоценозе обязательно наличие в качестве основного звена растительного сообщества (фитоценоз), обеспечивающего потенциальную бессмертность биогеоценоза за счет энергии, вырабатываемой растениями. Экосистемы могут не содержать фитоценоз.
Фитоценоз
Растительное сообщество, исторически сложившееся в результате сочетания взаимодействующих растений на однородном участке территории.
Его характеризуют :
- определенный видовой состав,
- жизненные формы,
- ярусность (надземная и подземная),
- обилие (частота встречаемости видов),
- размещение,
- аспект (внешний вид),
- жизненность,
- сезонные изменения,
- развитие (смена сообществ).
Ярусность (этажность)
Один из характерных признаков растительного сообщества, заключающийся как бы в поэтажном его разделении как в надземном, так и в подземном пространстве.
Надземная ярусность позволяет лучше использовать свет, а подземная - воду и минеральные вещества. Обычно в лесу можно выделить до пяти ярусов: верхний (первый) - высокие деревья, второй - невысокие деревья, третий - кустарники, четвертый - травы, пятый - мхи.
Подземная ярусность - зеркальное отражение надземной: глубже всех уходят корни деревьев, близ поверхности почвы расположены подземные части мхов.
По способу получения и использования питательных веществ все организмы делятся на автотрофы и гетеротрофы . В природе возникает непрерывный круговорот биогенных веществ, необходимых для жизни. Химические вещества извлекаются автотрофами из окружающей среды и через гетеротрофы вновь в нее возвращаются. Этот процесс принимает очень сложные формы. Каждый вид использует лишь часть содержащейся в органическом веществе энергии, доводя его распад до определенной стадии. Таким образом, в процессе эволюции в экологических системах сложились цепи и сети питания .
Большинство биогеоценозов имеют сходную трофическую структуру . Основу их составляют зеленые растения - продуценты. Обязательно присутствуют растительноядные и плотоядные животные: потребители органического вещества - консументы и разрушители органических остатков - редуценты .
Количество особей в пищевой цепи последовательно уменьшается, численность жертв больше численности их потребителей, так как в каждом звене пищевой цепи при каждом переносе энергии 80-90% ее теряется, рассеиваясь в виде теплоты. Поэтому число звеньев в цепи ограничено (3-5).
Видовое разнообразие биоценоза представлено всеми группами организмов - продуцентами, консументами и редуцентами.
Нарушение какого-либо звена в цепи питания вызывает нарушение биоценоза в целом. Например, вырубка леса приводит к изменению видового состава насекомых, птиц, а, следовательно, и зверей. На безлесном участке будут складываться другие цепи питания и сформируется другой биоценоз, что займет не один десяток лет.
Цепь питания (трофическая или пищевая )
Взаимосвязанные виды, последовательно извлекающие органическое вещество и энергию из исходного пищевого вещества; при этом каждое предыдущее звено цепи является пищей для последующего.
Цепи питания в каждом природном участке с более или менее однородными условиями существования составлены комплексами взаимосвязанных видов, питающимися друг другом и образующими самоподдерживающуюся систему, в которой осуществляется круговорот веществ и энергии.
Компоненты экосистемы:
- Продуценты - автотрофные организмы (в основном зеленые растения) - единственные производители органического вещества на Земле. Богатое энергией органическое вещество в процессе фотосинтеза синтезируется из бедных энергией неорганических веществ (Н 2 0 и С0 2).
- Консументы - растительноядные и плотоядные животные, потребители органического вещества. Консументы могут быть растительноядными, когда они непосредственно используют продуценты, или плотоядными, когда они питаются другими животными. В цепи питания они чаще всего могут иметь порядковый номер с I по IV .
- Редуценты - гетеротрофные микроорганизмы (бактерии) и грибы - разрушители органических остатков, деструкторы. Их еще называют санитарами Земли.
Трофический (пищевой) уровень - совокупность организмов, объединяемых типом питания. Представление о трофическом уровне позволяет понять динамику потока энергии в экосистеме.
- первый трофический уровень всегда занимают продуценты (растения),
- второй - консументы I порядка (растительноядные животные),
- третий - консументы II порядка - хищники, питающиеся растительноядными животными),
- четвертый - консументы III порядка (вторичные хищники).
Различают следующие виды пищевых цепей:
В пастбищной цепи (цепи выедания ) основным источником пищи служат зеленые растения. Например: трава -> насекомые -> земноводные -> змеи -> хищные птицы.
- детритные цепи (цепи разложения) начинаются с детрита - отмершей биомассы. Например: листовой опад -> дождевые черви -> бактерии. Особенностью детритных цепей является также то, что в них часто продукция растений не потребляется непосредственно растительноядными животными, а отмирает и минерализуется сапрофитами. Детритные цепи характерны также для экосистем океанических глубин, обитатели которых питаются мертвыми организмами, опустившимися вниз из верхних слоев воды.
Сложившиеся в процессе эволюции взаимоотношения между видами в экологических системах, при которых многие компоненты питаются разными объектами и сами служат пищей различным членам экосистемы. Упрощенно пищевую сеть можно представить как систему переплетающихся пищевых цепей .
Организмы разных пищевых цепей, получающие пищу через равное число звеньев этих цепей, находятся на одном трофическом уровне . В то же время разные популяции одного и того же вида, входящие в различные пищевые цепи, могут находиться на разных трофических уровнях . Соотношение различных трофических уровней в экосистеме можно изобразить графически в виде экологической пирамиды .
Экологическая пирамида
Способ графического отображения соотношения различных трофических уровней в экосистеме - бывает трех типов :
Пирамида численности отражает численность организмов на каждом трофическом уровне;
Пирамида биомасс отражает биомассу каждого трофического уровня;
Пирамида энергии показывает количество энергии, прошедшее через каждый трофический уровень в течение определенного промежутка времени.
Правило экологической пирамиды
Закономерность, отражающая прогрессивное уменьшение массы (энергии, числа особей) каждого последующего звена пищевой цепи.
Пирамида численности
Экологическая пирамида, отражающая число особей на каждом пищевом уровне. В пирамиде чисел не учитываются размеры и масса особей, продолжительность жизни, интенсивность обмена веществ, однако всегда прослеживается главная тенденция - уменьшение числа особей от звена к звену. Например, в степной экосистеме численность особей распределяется так: продуценты - 150000, травоядные консументы - 20000, плотоядные консументы - 9000 экз./ар. Биоценоз луга характеризуется следующей численностью особей на площади 4000 м 2: продуценты - 5 842 424, растительноядные консументы I порядка - 708 624, плотоядные консументы II порядка - 35 490, плотоядные консументы III порядка - 3.
Пирамида биомасс
Закономерность, согласно которой количество растительного вещества, служащего основой цепи питания (продуцентов), примерно в 10 раз больше, чем масса растительноядных животных (консументов I порядка), а масса растительноядных животных в 10 раз больше, чем плотоядных (консументов II порядка), т. е. каждый последующий пищевой уровень имеет массу в 10 раз меньшую, чем предыдущий. В среднем из 1000 кг растений образуется 100 кг тела травоядных животных. Хищники, поедающие травоядных, могут построить 10 кг своей биомассы, вторичные хищники - 1 кг.
Пирамида энергии
выражает закономерность, согласно которой поток энергии постепенно уменьшается и обесценивается при переходе от звена к звену в цепи питания. Так, в биоценозе озера зеленые растения - продуценты - создают биомассу, содержащую 295,3 кДж/см 2 , консументы I порядка, потребляя биомассу растений, создают свою биомассу, содержащую 29,4 кДж/см 2 ; консументы II порядка, используя в пищу консументов I порядка, создают свою биомассу, содержащую 5,46 кДж/см 2 . Потеря энергии при переходе от консументов I порядка к консументам II порядка, если это теплокровные животные, увеличивается. Это объясняется тем, что у данных животных много энергии уходит не только на построение своей биомассы, но и на поддержание постоянства температуры тела. Если сравнить выращивание теленка и окуня, то одинаковое количество затраченной пищевой энергии даст 7 кг говядины и лишь 1 кг рыбы, так как теленок питается травой, а окунь-хищник - рыбой.
Таким образом , первые два типа пирамид имеют ряд существенных недостатков:
Пирамида биомасс отражает состояние экосистемы на момент отбора пробы и, следовательно, показывает соотношение биомассы в данный момент и не отражает продуктивность каждого трофического уровня (т. е. его способность образовывать биомассу в течение определенного промежутка времени). Поэтому в том случае, когда в число продуцентов входят быстрорастущие виды, пирамида биомасс может оказаться перевернутой.
Пирамида энергии позволяет сравнить продуктивность различных трофических уровней, поскольку учитывает фактор времени. Кроме того, она учитывает разницу в энергетической ценности различных веществ (например, 1 г жира дает почти в два раза больше энергии, чем 1 г глюкозы). Поэтому пирамида энергии всегда суживается кверху и никогда не бывает перевернутой.
Экологическая пластичность
Степень выносливости организмов или их сообществ (биоценозов) к воздействию факторов среды. Экологически пластичные виды имеют широкую норму реакции , т. е. широко приспособлены к разной среде обитания (рыбы колюшка и угорь, некоторые простейшие живут как в пресных, так и в соленых водах). Узкоспециализированные виды могут существовать лишь в определенной среде: морские животные и водоросли - в соленой воде, речные рыбы и растения лотос, кувшинка, ряска обитают только в пресной воде.
В целом экосистема (биогеоценоз) характеризуется следующими показателями :
Видовым разнообразием,
Плотностью видовых популяций,
Биомассой.
Биомасса
Общее количество органического вещества всех особей биоценоза или вида с заключенной в нем энергией. Биомассу выражают обычно в единицах массы в пересчете на сухое вещество единицы площади или объема. Биомассу можно определить отдельно для животных, растений или отдельных видов. Так, биомасса грибов в почве составляет 0,05-0,35 т/га, водорослей - 0,06-0,5, корней высших растений - 3,0-5,0, дождевых червей - 0,2-0,5, позвоночных животных - 0,001-0,015 т/га.
В биогеоценозах различают первичную и вторичную биологическую продуктивность :
ü Первичная биологическая продуктивность биоценозов - общая суммарная продуктивность фотосинтеза, представляющая собой результат деятельности автотрофов - зеленых растений, например, сосновый лес 20- 30-летнего возраста за год производит 37,8 т/га биомассы.
ü Вторичная биологическая продуктивность биоценозов - общая суммарная продуктивность гетеротрофных организмов (консументов), которая образуется за счет использования веществ и энергии, накопленных продуцентами.
Популяции. Структура и динамика численности.
Каждый вид на Земле занимает определенный ареал , так как он способен существовать лишь в определенных условиях среды. Однако условия обитания в рамках ареала одного вида могут существенно отличаться, что приводит к распаду вида на элементарные группировки особей - популяции.
Популяция
Совокупность особей одного вида, занимающих обособленную территорию в пределах ареала вида (с относительно однородными условиями обитания), свободно скрещивающихся друг с другом (имеющих общий генофонд) и изолированных от других популяций данного вида, обладающих всеми необходимыми условиями для поддержания своей стабильности длительное время в меняющихся условиях среды. Важнейшими характеристиками популяции являются ее структура (возрастной, половой состав) и динамика численности.
Под демографической структурой популяции понимают ее половой и возрастной состав.
Пространственная структура популяции - это особенности размещения особей популяции в пространстве.
Возрастная структура популяции связана с соотношением особей различных возрастов в популяции. Особи одного возраста объединяют в когорты - возрастные группы.
В возрастной структуре популяций растений выделяют следующие периоды :
Латентный - состояние семени;
Прегенеративный (включает состояния проростка, ювенильного растения, имматурного и виргинильного растений);
Генеративный (обычно подразделяется на три подпериода - молодые, зрелые и старые генеративные особи);
Постгенеративный (включает состояния субсенильного, сенильного растений и фазу отмирания).
Принадлежность к определенному возрастному состоянию определяется по биологическому возрасту - степени выраженности определенных морфологических (например, степень расчлененности сложного листа) и физиологических (например, способность дать потомство) признаков.
В популяциях животных также можно выделить различные возрастные стадии . Например, насекомые, развивающиеся с полным метаморфозом, проходят стадии:
Личинки,
Куколки,
Имаго (взрослого насекомого).
Характер возрастной структуры популяции зависит от типа кривой выживания, свойственной данной популяции.
Кривая выживания отражает уровень смертности в различных возрастных группах и представляет собой снижающуюся линию:
- Если уровень смертности не зависит от возраста особей, отмирание особей происходит в данном типе равномерно, коэффициент смертности остается постоянным на протяжении всей жизни (тип I ). Такая кривая выживания свойственна видам, развитие которых происходит без метаморфоза при достаточной устойчивости рождающегося потомства. Этот тип принято называть типом гидры - для нее свойственна кривая выживания, приближающаяся к прямой линии.
- У видов, для которых роль внешних факторов в смертности невелика, кривая выживания характеризуется небольшим понижением до определенного возраста, после которого происходит резкое падение вследствие естественной (физиологической) смертности (тип II ). Близкий к этому типу характер кривой выживания свойствен человеку (хотя кривая выживания человека несколько более пологая и является чем-то средним между типами I и II). Этот тип носит название типа дрозофилы : именно его демонстрируют дрозофилы в лабораторных условиях (не поедаемые хищниками).
- Для очень многих видов характерна высокая смертность на ранних стадиях онтогенеза. У таких видов кривая выживания характеризуется резким падением в области младших возрастов. Особи, пережившие «критический» возраст, демонстрируют низкую смертность и доживают до старших возрастов. Тип носит название типа устрицы (тип III ).
Половая структура популяции
Соотношение полов имеет прямое отношение к воспроизводству популяции и ее устойчивости.
Выделяют первичное, вторичное и третичное соотношение полов в популяции:
- Первичное соотношение полов определяется генетическими механизмами - равномерностью расхождения половых хромосом. Например, у человека XY-хромосомы определяют развитие мужского пола, а XX - женского. В этом случае первичное соотношение полов 1:1, т. е. равновероятно.
- Вторичное соотношение полов - это соотношение полов на момент рождения (среди новорожденных). Оно может существенно отличаться от первичного по целому ряду причин: избирательность яйцеклеток к сперматозоидам, несущим Х- или Y-хромосому, неодинаковой способностью таких сперматозоидов к оплодотворению, различными внешними факторами. Например, зоологами описано влияние температуры на вторичное соотношение полов у рептилий. Аналогичная закономерность характерна и для некоторых насекомых. Так, у муравьев оплодотворение обеспечивается при температуре выше 20 °С, а при более низких температурах откладываются неоплодотворенные яйца. Из последних вылупляются самцы, а из оплодотворенных - преимущественно самки.
- Третичное соотношение полов - соотношение полов среди взрослых животных.
Пространственная структура популяции отражает характер размещения особей в пространстве.
Выделяют три основных типа распределения особей в пространстве:
- единообразное или равномерное (особи размещены в пространстве равномерно, на одинаковых расстояниях друг от друга); встречается в природе редко и чаще всего вызвано острой внутривидовой конкуренцией (например, у хищных рыб);
- конгрегационное или мозаичное («пятнистое», особи размещаются в обособленных скоплениях); встречается намного чаше. Оно связано с особенностями микросреды или поведения животных;
- случайное или диффузное (особи распределены в пространстве случайным образом) - можно наблюдать только в однородной среде и только у видов, которые не обнаруживают никакого стремления к объединению в группы (например, у жука в муке).
Численность популяции обозначается буквой N. Отношение прироста N к единице времени dN / dt выражает мгновенную скорость изменения численности популяции, т. е. изменение численности в момент времени t. Прирост популяции зависит от двух факторов - рождаемости и смертности при условии отсутствия эмиграции и иммиграции (такая популяция называется изолированной). Разность рождаемости b и смертности d и представляет собой коэффициент прироста изолированной популяции :
Устойчивость популяции
Это ее способность находиться в состоянии динамического (т. е. подвижного, изменяющегося) равновесия со средой: изменяются условия среды - изменяется и популяция. Одним из важнейших условий устойчивости является внутреннее разнообразие. Применительно к популяции это механизмы поддержания определенной плотности популяции.
Выделяют три типа зависимости численности популяции от ее плотности .
Первый тип (I) - самый распространенный, характеризуется уменьшением роста популяции при увеличении ее плотности, что обеспечивается различными механизмами. Например, для многих видов птиц характерны снижение рождаемости (плодовитости) при увеличении плотности популяции; увеличение смертности, снижение сопротивляемости организмов при повышенной плотности популяции; изменение возраста наступления половой зрелости в зависимости от плотности популяции.
Третий тип ( III ) характерен для популяций, в которых отмечается «эффект группы», т. е. определенная оптимальная плотность популяции способствует лучшему выживанию, развитию, жизнедеятельности всех особей, что присуще большинству групповых и социальных животных. Например, для возобновления популяций разнополых животных как минимум необходима плотность, обеспечивающая достаточную вероятность встречи самца и самки.
Тематические задания
А1. Биогеоценоз образован
1) растениями и животными
2) животными и бактериями
3) растениями, животными, бактериями
4) территорией и организмами
А2. Потребителями органического вещества в лесном биогеоценозе являются
1) ели и березы
2) грибы и черви
3) зайцы и белки
4) бактерии и вирусы
А3. Продуцентами в озере являются
2) головастики
А4. Процесс саморегуляции в биогеоценозе влияет на
1) соотношение полов в популяциях разных видов
2) численность мутаций, возникающих в популяциях
3) соотношение хищник – жертва
4) внутривидовую конкуренцию
А5. Одним из условий устойчивости экосистемы может служить
1) ее способность к изменениям
2) разнообразие видов
3) колебания численности видов
4) стабильность генофонда в популяциях
А6. К редуцентам относятся
2) лишайники
4) папоротники
А7. Если общая масса полученной потребителем 2-го порядка равна 10 кг, то какова была совокупная масса продуцентов, ставших источником пищи для данного потребителя?
А8. Укажите детритную пищевую цепь
1) муха – паук – воробей – бактерии
2) клевер – ястреб – шмель – мышь
3) рожь – синица – кошка – бактерии
4) комар – воробей – ястреб – черви
А9. Исходным источником энергии в биоценозе является энергия
1) органических соединений
2) неорганических соединений
4) хемосинтеза
1) зайцами
2) пчелами
3) дроздами-рябинниками
4) волками
А11. В одной экосистеме можно встретить дуб и
1) суслика
3) жаворонка
4) синий василек
А12. Сети питания – это:
1) связи между родителями и потомством
2) родственные (генетические) связи
3) обмен веществ в клетках организма
4) пути передачи веществ и энергии в экосистеме
А13. Экологическая пирамида чисел отражает:
1) соотношение биомасс на каждом трофическом уровне
2) соотношение масс отдельного организма на разных трофических уровнях
3) структуру пищевой цепи
4) разнообразие видов на разных трофических уровнях
Перенос энергии пищи от его источника – автотрофов (растений) – через ряд организмов, происходящий путём поедания одних организмов другими, называется пищевой цепью .
При каждом очередном переносе большая часть потенциальной энергии (80÷90%) теряется, переходя в тепло. Поэтому чем короче пищевая цепь (чем ближе организм к её началу – солнечной энергии), тем больше количество энергии, доступной для популяции.
Пищевые цепи можно разделить на два основных типа: пастбищная цепь , которая начинается с зелёного растения и идёт далее к пасущимся растительноядным животным и к их хищникам, идетритная цепь , которая от мёртвого органического вещества идёт к микроорганизмам, а затем к детритофагам и к их хищникам. Пищевые цепи не изолированы одна от другой, а тесно переплетаются друг с другом, образуя так называемыепищевые сети .
Пастбищная
Солнечный Растительноядные Хищники
Детритная
Потребители детрита Хищники
Наиболее упрощенные пастбищная и детритная пищевые цепи объединены в пищевую сеть в виде Y-образной или двухканальной диаграммы потока энергии .
Величины тех частей энергии чистой продукции, которые текут по двум путям, различны в экосистемах разного типа и часто варьируют по сезонам или по годам в одной и той же экосистеме. На некоторых мелководьях и на интенсивно используемых пастбищах и в степях по пастбищной цепи может идти 50% и более чистой продукции. Напротив, прибрежные марши, океаны, леса, да и большинство природных экосистем, функционируют как детритные системы; в них 90% и более процентов автотрофной продукции потребляется гетеротрофами только после того, как листья, стебли и другие части растений отмирают, подвергаются «переработке», превращаясь в диспергированное или растворённое органическое вещество, поступающее в воду, донные осадки и почву. Такое отсроченное потребление увеличивает структурную сложность, а также накопительную и буферную ёмкость экосистем.
Тесная связь пастбищной и детритной пищевых цепей приводит к тому, что при изменении уровня энергетического воздействия извне на экосистему быстро происходит переключение потоков между каналами, что позволяет поддерживать устойчивость экосистем. Не вся пища, съеденная пасущимися животными, усваивается: часть её, например через фекалии, уходит в детритную цепь.
Степень влияния травоядных животных на сообщество зависит не только от количества ассимилированной ими энергии пищи, но и от скорости изъятия живых растений. Прямое изъятие травоядными животными или человеком более 30-50% годового прироста наземной растительности уменьшает способность экосистемы сопротивляться стрессу. Перевыпас скота был одной из причин упадка многих цивилизаций. «Недовыпас » также может оказаться вредным. Если прямое потребление живых растений совершенно отсутствует, то детрит может накапливаться быстрее, чем идёт его разложение микроорганизмами. Это замедляет круговорот минеральных веществ, и, кроме того, система может стать пожароопасной.
В сложных природных сообществах организмы, получающие свою энергию от Солнца через одинаковое число ступеней, считаются принадлежащими к одному трофическому уровню . Так, зелёные растения занимают первый трофический уровень (уровень продуцентов), травоядные – второй (уровень первичных консументов), первичные хищники, поедающие травоядных, – третий (уровень вторичных консументов), а вторичные хищники – четвёртый (уровень третичных консументов). Эта трофическая классификация относится к функциям, а не к видам как таковым. Популяция данного вида может занимать один или несколько трофических уровней, смотря по тому, какие источники энергии она использует. Поток энергии через трофический уровень равен общей ассимиляции (А ) на этом уровне, которая в свою очередь равна продукции (Р ) биомассы плюс дыхание (R ):
A =P +R .
При переносе энергии между трофическими уровнями часть потенциальной энергии теряется. Прежде всего, растение фиксирует лишь малую долю поступающей энергии солнечного света (около 1%). Поэтому число консументов (например, людей), которые могут прожить при данном выходе первичной продукции, сильно зависит от длины пищевой цепи; переход к каждому следующему звену в нашей традиционной сельскохозяйственной пищевой цепи уменьшает доступную энергию примерно на порядок величины (т.е. в 10 раз). Поэтому если в рационе увеличивается содержание мяса, то уменьшается число людей, которых можно прокормить.
Эффективность i-го трофического уровня принято оценивать, как отношение A i /A i-1 , гдеA i – ассимиляцияi -ым трофическим уровнем. Для первого (автотрофного) трофического уровня она составляет 1-5%, для последующих – 10-20%.
Может озадачить низкая эффективность природных экосистем в сравнении с высокими КПД электромоторов и других двигателей. Но на самом деле, долгоживущие, крупномасштабные экосистемы нельзя приравнивать в этом отношении к недолговечным механическим системам. Во-первых, в живых системах много «горючего» затрачивается на «ремонт» и самоподдержание, а при расчете КПД двигателей не учитываются амортизация и расходы энергии на ремонт. Во-вторых, в определенных условиях быстрый рост, который повышает потребление энергии, может иметь большее значение для выживания, чем максимальная эффективность использования энергии пищи или горючего.
Для экосистем важно понимать, что любое повышение их эффективности искусственным путём обернется увеличением затрат на ее поддержание. Всегда наступает такой предел, после которого выигрыш от роста эффективности сводится на нет ростом расходов, не говоря уже о том, что система может войти в опасное колебательное состояние, грозящее разрушением. Индустриализованные экосистемы, возможно, уже достигли такой стадии, когда увеличение расходов приводит к все меньшей отдаче.
Пищевая цепь состоит из организмов разных видов. В то же время организмы одного вида могут входить в состав разных пищевых цепей. Поэтому цепи питания переплетаются, образуя сложные пищевые сети, охватывающие все экосистемы планеты.[ ...]
Пищевая (трофическая) цепь - это перенос энергии от ее источника - продуцентов - через ряд организмов. Пищевые цепи можно разделить на два основных типа: пастбищная цепь, которая начинается с зеленого растения и идет далее к пасущимся растительноядным животным и к хищникам, и детритная цепь (от латинского истертый), которая начинается от продуктов распада мертвого органического вещества. В формировании этой цепи решающую роль играют различные микроорганизмы, которые питаются мертвым органическим веществом и минерализуют его, вновь превращая в простейшие неорганические соединения. Пищевые цепи не изолированы одна от другой, а тесно переплетаются друг с другом. Часто животное, потребляющее живое органическое вещество, поедает и микробов, потребляющих в пищу неживое органическое вещество. Таким образом, пути потребления пищи разветвляются, образуя так называемые пищевые сети.[ ...]
Пищевая сеть - сложное переплетение в сообществе пищевых цепей.[ ...]
Пищевые сети образуются потому, что практически любой член какой-либо пищевой цепи одновременно является звеном и в другой пищевой цепи: он потребляет и его потребляют несколько видов других организмов. Так, в пище лугового волка - койота насчитывают до 14 тыс. видов животных и растений. Вероятно, таков же порядок числа видов, участвующих в поедании, разложении и деструкции веществ трупа койота.[ ...]
Пищевые цепи и трофические уровни. Прослеживая пищевые взаимоотношения между членами биоценоза («кто кого и сколько поедает»), можно построить пищевые цепи питания различных организмов. Примером длинной пищевой цепи может служить последовательность обитателей арктического моря: «микроводоросли (фитопланктон) -> мелкие растительноядные ракообразные (зоопланктон) - плотоядные планктонофаги (черви, ракообразные, моллюски, иглокожие) -> рыбы (возможны 2-3 звена последовательности хищных рыб) -> тюлени -> белый медведь». Цепи наземных экосистем обычно короче. Пищевая цепь, как правило, искусственно выделяется из реально существующей пищевой сети - сплетения многих цепей питания.[ ...]
Пищевая сеть - это сложная сеть пищевых взаимоотношений.[ ...]
Пищевые цепи подразумевают линейный поток ресурсов от одного трофического уровня к следующему (рис. 22.1, а). В такой конструкции взаимодействия между видами просты. Однако никакая система потоков ресурсов в БЭ не следует этой простой структуре; они гораздо больше напоминают сетевую структуру (рис. 22.1, Ь). Здесь виды на одном трофическом уровне питаются несколькими видами на следующем, более низком уровне и широко распространена всеядность (рис. 22.1, с). Наконец, полностью определенная пищевая сеть может продемонстрировать различные особенности: множество трофических уровней, хищничество и всеядность (рис. 22.1, [ ...]
Множество пищевых цепей, переплетаясь в биоценозах и экосистемах, образуют пищевые сети. Если общую цепь питания изобразить в виде строительных блоков, условно представляющих собой количественное соотношение усваиваемой на каждом этапе энергии, и сложить их друг на друга, получится пирамида. Ее называют экологической пирамидой энергий (рис.5).[ ...]
Диаграммы пищевых цепей и пищевых сетей. Точки обозначают виды, линии обозначают взаимодействия. Более высокие виды являются хищниками по отношению к более низким, поэтому ресурсы текут снизу вверх.[ ...]
В первом типе пищевой сети поток энергии идет от растений к растительноядным животным, а далее к консументам более высокого порядка. Это сеть выедания, или пастбищная сеть. Вне зависимости от величины биоценоза и места обитания растительноядные животные (наземные, водные, почвенные) пасутся, выедают зеленые растения и передают энергию на следующие уровни (рис. 96).[ ...]
В сообществах пищевые цепи сложным образом переплетаются и образуют пищевые сети. В состав пищи каждого вида входит обычно не один, а несколько видов, каждый из которых в свою очередь может служить пищей нескольким видам. С одной стороны, каждый трофический уровень представлен многими популяциями разных видов, с другой стороны, многие популяции принадлежат сразу к нескольким трофическим уровням. В результате благодаря сложности пищевых связей выпадение какого-то одного вида часто не нарушает равновесия в экосистеме.[ ...]
[ ...]
Эта схема не только иллюстрирует переплетение пищевых связей и показывает три трофических уровня, но и выявляет тот факт, что некоторые организмы занимают промежуточное положение в системе трех основных трофических уровней. Так, строящие ловчую сеть личинки ручейника питаются растениями и животными, занимая промежуточное положение между первичными и вторичными консументами.[ ...]
Первичным источником пищевых ресурсов человека были те экосистемы, в которых он мог существовать. Способами добывания пищи были собирательство и охота, причем по мере развития изготовления и использования все более совершенных орудий увеличивалась доля охотничьей добычи, значит, и доля мяса, то есть полноценных белков, в рационе. Способность организовывать большие устойчивые коллективы, развитие речи, позволяющей организовывать сложное согласованное поведение множества людей, сделали человека «суперхищником», занявшим верхнюю позицию в пищевых сетях тех экосистем, которые он осваивал по мере расселения по Земле. Так, единственным врагом мамонта был человек, который, вместе с отступлением ледника и изменением климата стал одной из причин гибели этих северных слонов как вида.[ ...]
[ ...]
На основании изучения 14 пищевых сетей в сообществах Коэн обнаружил удивительное постоянство отношения числа «типов» жертвы к числу «типов» хищников, составлявшее примерно 3: 4. Дальнейшие данные, подтверждающие это соотношение, приводят Брайанд и Коэн , исследовавшие 62 аналогичные сети. График такой пропорциональности имеет тангенс угла наклона менее 1 как во флуктуирующих, так и в постоянных средах. Использование «типов» организмов, а не подлинных видов обычно дает не вполне объективные результаты , однако, хотя полученное при этом отношение жертва/хищник, возможно, занижено, его постоянство примечательно.[ ...]
В БЭ многие (но определенно не все) пищевые сети имеют большое количество первичных производителей, меньше потребителей и совсем немного высших хищников, что придает сети форму, приведенную на рис. 22.1, Ь. Всеядные в этих системах могут быть редки, в то время как редуценты находятся в изобилии. Модели пищевых сетей обеспечили потенциальную основу для плодотворного анализа потоков ресурсов как в БЭ, так и в ПЭ. Сложности возникают, однако, когда пытаются количественно определить потоки ресурсов и подвергнуть структуру сети и свойства стабильности математическому анализу. Оказывается, что многие из необходимых данных трудно выявить с определенностью, особенно в том, что касается организмов, которые функционируют более чем на одном трофическом уровне. Эго свойство создает не основную сложность исследований потоков ресурсов, но оно серьезно усложняет анализ стабильности. Утверждение, что более сложные системы являются более стабильными - поскольку разрушение одного вида или путей потоков просто переводит энергию и ресурсы на другие пути, а не блокирует путь для всего потока энергии или ресурсов - пока горячо обсуждается.[ ...]
Анализ большого числа промышленных пищевых сетей может, таким образом, выявить характеристики, не показанные в других подходах. В проекте экосистемы на рис. 22.5, например, сетевой анализ может отразить отсутствующий сектор или тип промышленной деятельности, который способен увеличить связанность. Эти темы дают богатую область для детальных исследований.[ ...]
Внутри каждой экосистемы трофические сети имеют хорошо выраженную структуру, которая характеризуется природой и количеством организмов, представленных на каждом уровне различных пищевых цепей. Для изучения взаимоотношений между организмами в экосистеме и для их графического изображения обычно используют не схемы пищевых сетей, а экологические пирамиды. Экологические пирамиды выражают трофическую структуру экосистемы в геометрической форме.[ ...]
Некоторый интерес представляет длина пищевых цепей. Ясно, что уменьшение доступной энергии при переходе к каждому последующему звену ограничивает длину пищевых цепей. Однако доступность энергии, видимо, не единственный фактор, поскольку длинные пищевые цепи часто встречаются в неплодородных системах, например в олиготрофных озерах, а короткие - в очень продуктивных, или эвтрофных, системах. Быстрое продуцирование питательного растительного материала может стимулировать быстрое выедание, в результате чего поток энергии концентрируется на первых двух-трех трофических уровнях. Эвтрофикация озер также изменяет состав планктонной пищевой сети «фитопланктон-крупный зоопланктон-хищная рыба», превращая его в микробно-детритную микрозоопланктонную систему, не столь способствующую поддержанию спортивного рыболовства.[ ...]
При неизменном энергетическом потоке в пищевой сети, или цепи, более мелкие наземные организмы с высоким удельным метаболизмом создают относительно меньшую биомассу, чем крупные1. Значительная часть энергии уходит на поддержание обмена веществ. Это правило «метаболизм и размеры особей», или правило Ю. Одума, обычно не реализуется в водных биоценозах при учете реальных условий обитания в них (в идеальных условиях оно имеет всеобщее значение). Связано это с тем, что мелкие водные организмы в значительной мере поддерживают свой обмен веществ за счет внешней энергии непосредственно окружающей их среды.[ ...]
Почвенная микрофлора имеет хорошо развитую пищевую сеть и мощный компенсационный механизм, основанный на функциональной взаимозаменяемости одних видов другими. Кроме того, благодаря лабильному ферментативному аппарату многие виды могут легко переключаться с одного питательного субстрата на другой, обеспечивая тем самым стабильность экосистемы. Это существенно усложняет оценку воздействия на нее различных антропогенных факторов и требует использование интегральных показателей.[ ...]
[ ...]
Прежде всего рандомизированно составленные пищевые сети часто содержат биологически бессмысленные элементы (например, петли такого типа: А поедает В, В поедает С, С поедает А). Анализ «осмысленно» построенных сетей (Lawlor, 1978; Pimm, 1979а) показывает, что (а) они устойчивее рассмотренных и (б) нет такого резкого перехода к неустойчивости (по сравнению с приведенным выше неравенством), хотя устойчивость по-прежнему падает с ростом сложности.[ ...]
| 21.2 |  |
Безусловно, да, если не в составе биогеоценозов - низших ступеней иерархии экосистем,- то уж, во всяком случае, в рамках биосферы. Люди из этих сетей получают пищу (агроценозы- видоизмененные экосистемы с природной основой). Только из «дикой» природы люди извлекают топливо - энергию, основные рыбные ресурсы, другие «дары природы». Мечта В. И. Вернадского о полной автотрофности человечества пока остается иррациональной мечтой1 - эволюция необратима (правило Л. Доло), как и исторический процесс. Без подлинных автотрофов, в основном растений, человек не может существовать как гетеротрофный организм. Наконец, если бы он физически не был включен в пищевые сети природы, то его тело после смерти не подвергалось бы разрушению организмами-редуцентами, и Земля была бы завалена несгнившими трупами. Тезис о раздельности человека и природных пищевых цепей основан на недоразумении и явно ошибочен.[ ...]
В гл. 17 анализируются способы объединения различных групп консументов и их пищи в сеть взаимодействующих элементов, по которой происходит передача вещества и энергии. В гл. 21 мы возвратимся к этой теме и рассмотрим влияние структуры пищевой сети на динамику сообществ в целом, обратив особое внимание на особенности их структуры, способствующие стабильности.[ ...]
Четырех примеров будет достаточно, чтобы проиллюстрировать основные особенности пищевых цепей, пищевых сетей и трофических уровней. Первый пример - регион Крайнего Севера, называемый тундрой, где обитает сравнительно немного видов организмов, успешно приспособившихся к низким температурам. Поэтому пищевые цепи и пищевые сети здесь относительно просты. Один из основателей современной экологии, британский эколог Ч. Элтон, поняв это, уже в 20-30-х годах нашего века занялся изучением арктических земель. Он одним из первых четко обрисовал принципы и концепции, связанные с пищевыми цепями (Elton, 1927). Растения тундры - лишайник («олений мох») С1а donia, злаки, осоки и карликовые ивы составляют пищу оленя карибу в североамериканской тундре и его экологического аналога в тундре Старого Света - северного оленя. Эти животные в свою очередь служат пищей волкам и человеку. Тундровые растения поедают также лемминги - пушистые короткохвостые грызуны, напоминающие медведя в миниатюре, и тундряные куропатки. Всю долгую зиму и все короткое лето песцы и полярные совы питаются в основном леммингами. Любое значительное изменение численности леммингов отражается и на других трофических уровнях, так как других источников пищи мало. Вот почему численность некоторых групп арктических организмов сильно колеблется от сверхизобилия до почти полного исчезновения. Подобное часто случалось в человеческом обществе, если оно зависело от одного или нескольких немногих источников пищи (вспомним «картофельный голод» в Ирландии1).[ ...]
Одно из следствий гипотезы устойчивости, которое в прин-пице можно проверить, это то, что в средах с менее предсказуемым поведением пищевые цепи должны быть короче, поскольку в них, по-видимому, сохраняются лишь наиболее упругие пищевые сети, а у коротких цепей упругость выше. Брай-анд (Briand, 1983) разделил 40 пищевых сетей (по собранным им данным) на связанные с изменчивой (позиции 1-28 в табл. 21.2) и постоянной (позиции 29-40) средами. Достоверных различий в средней длине максимальных пищевых цепей между этими группами не обнаружилось: число трофических уровней составило 3,66 и 3,60 соответственно (рис. 21.9). Эти положения еще нуждаются в критической проверке.[ ...]
Кроме того, результаты моделирования становятся иными, когда учитывается, что популяции консументов испытывают влияние со стороны пищевых ресурсов, а те от воздействия консументов не зависят (¡3,/Х), 3(/ = 0: так называемая «система, регулируемая донором»), В пищевой сети подобного типа устойчивость либо не зависит от сложности, либо повышается вместе с ней (DeAngelis, 1975). На практике единственной группой организмов, которая обычно удовлетворяет этому условию, являются детритофаги.[ ...]
Однако такая строгая картина перехода энергии с уровня на уровень не совсем реальна, поскольку трофические цепи экосистем сложно переплетаются, образуя трофические сети. Например, явление «трофического каскада», когда в результате хищничества происходит изменение плотности, биомассы или продуктивности популяции, сообщества или трофического уровня по более чем одной линии пищевой сети (Пейс и др., 1999). П. Митчел (2001) приводит такой пример: морские выдры питаются морскими ежами, которые едят бурые водоросли, уничтожение охотниками выдр привело к уничтожению бурых водорослей вследствие роста популяции ежей. Когда запретили охоту на выдр, водоросли стали возвращаться на свои места обитания.[ ...]
Зеленые растения преобразуют энергию фотонов солнечного света в энергию химических связей сложных органических соединений, которые продолжают свой путь по разветвленным пищевым сетям природных экосистем. Однако в некоторых местах (например, на болотах, в устьях рек и морях) часть органических растительных веществ, попав на дно, покрывается песком раньше, чем станет пищей для животных или микроорганизмов. При наличии определенной температуры и давления грунтовых пород в течение тысяч и миллионов лет из органических веществ образуются уголь, нефть и прочее ископаемое топливо или, по выражению В. И. Вернадского, «живое вещество уходит в геологию».[ ...]
Примеры пишевых цепей: растения - растительноядные животные -хищник; злак-полевая мышь-лиса; кормовые растения - корова - человек. Как правило, каждый вид питается не одним-единственным видом. Поэтому пищевые цепи переплетаются, образуя пищевую сеть. Чем сильнее организмы связаны между собой пищевыми сетями и другими взаимодействиями, тем устойчивее сообщество против возможных нарушений. Естественные, ненарушенные экосистемы стремятся к равновесию. Состояние равновесия основано на взаимодействии биотических и абиотических факторов среды.[ ...]
Например, уничтожение ядохимикатами хозяйственно значимых вредителей в лесах, отстрел части популяций животных, вылов отдельных видов промысловых рыб - это частичные помехи, поскольку они влияют лишь на отдельные звенья пищевых цепей, не затрагивая пищевых сетей в целом. Чем сложнее пищевая сеть, структура экосистемы, тем значимость таких помех меньше, и наоборот. В то же время выброс и сброс в атмосферу или воду химических ксенобиотиков, например оксидов серы, азота, углеводородов, соединений фтора, хлора, тяжелых металлов, радикально меняет качество среды, создает помехи на уровне продуцентов в целом, а значит, и ведет к полной деградации экосистемы: так как погибает основной трофический уровень - продуценты.[ ...]
Энергозависимая пропускная способность = (/гЛ -)/к В. Энергетическая схема примитивной системы в Уганде. Г. Энергетическая схема сельского хозяйства в Индии, где главным источником энергии служит свет, но поток энергии через скот н зерноаые регулируется человеком. Д. Энергетическая сеть высокомеханизированного сельского хозяйства. Высокие урожаи основаны на значительном вложении энергии путем использования ископаемого топлива, за счет которого выполняется работа, ранее производившаяся человеком и животными; при этом выпадает пищевая сеть животных и растений, которых приходилось «кормить» в двух предыдущих системах.[ ...]
Был предпринят целый ряд попыток проанализировать математически зависимость между сложностью сообщества и его устойчивостью, в большинстве из которых авторы пришли к примерно одинаковым выводам. Обзор таких публикаций дал Мей (May, 1981). В качестве примера рассмотрим его работу (May, 1972), демонстрирующую как сам метод, так и его недостатки. На каждый вид влияли его взаимодействия со всеми остальными видами; количественно влияние плотности вида / на рост численности i оценивалось показателем р. При полном отсутствии влияния равен нулю, у двух конкурирующих видов Рц и Pji отрицательны, в случае хищника (¿) и жертвы (/) Ру положителен, a jjji - отрицателен.[ ...]
Кислотные осадки вызывают летальные последствия для жизни в реках и водоемах. Многие озера Скандинавии и восточной части Северной Америки оказались настолько закисле-ны, что рыба не может не только нереститься в них, но и просто выжить. В 70-е годы в половине озер указанных регионов рыба полностью исчезла. Наиболее опасно подкисление океанических мелководий, ведущее к невозможности размножения многих морских беспозвоночных животных, что может вызвать разрыв пищевых сетей и глубоко нарушить экологическое равновесие в Мировом океане.[ ...]
Модели контролируемых донором взаимодействий по ряду признаков отличаются от традиционных моделей взаимодействий типа хищник-жертва Лотки-Воль-терры (гл. 10). Одно из важных отличий состоит в том, что как полагают, взаимодействующие группы видов, для которых характерна контролируемая донором динамика, особенно устойчивы и, далее, что эта устойчивость фактически не зависит от увеличения видового разнообразия и сложности пищевой сети или даже возрастает. Эта ситуация совершенно противоположна той, в которой применима модель Лотки-Вольтерры. Подробнее эти важные вопросы, касающиеся сложности пищевой сети и устойчивости сообщества, мы обсудим в гл. 21.